
Who made the Aurignacian?
Sat, May 30 2009 03:40 | Palaeoanthropology | Permalink

For most of the last century, the prevalent view among archaeologists was that Neandertals only made Mousterian tools. However, the discovery of the St Césaire 1 Neandertal skeleton and the Neandertal remains from Arcy-sur-Cure with Châtelperronian industry put paid to this idea. Châtelperronian tools show a mix of features otherwise found in the Mousterian and Aurignacian industries. What was particularly surprising about the Châtelperronian culture was not only the lithics but also the manufacture of bone tools and personal ornaments. At the site of Arcy-sur-Cure archaeologists found pierced teeth, ivory, shell, and bone in the Châtelperronian layers. The Szeletian industry of central Europe and the Uluzzian industry of Italy may also be related to the Châtelperronian.
The Châtelperronian has shown that Neandertals were more skilful than previously thought and has opened up the possibility that they may have been the authors of the Aurignacian. At Vindija Cave in Croatia Neandertal remains were speculated to be associated with an Aurignacian-like assemblage (Smith et al. 1999). The skeletal fragments from this site show clear Neandertal affinities. However, problems with stratigraphic control during excavations, as well as evidence of cryoturbation and bioturbation mean that the Neandertal-Aurignacian association is questionable.
However, many of the best associations of the Aurignacian with modern humans are equally problematic. At Bacho Kiro Cave in Bulgaria a "proto-Aurignacian" culture has been associated with some fragmentary human remains (Kozłowski 1982). However, dates from the "proto-Aurignacian" layer span over thousands of years suggesting (1) a very long accumulation of sediment, (2) contamination or (3) incorrect context. Moreover, the fragmentary nature of the remains has meant that a taxonomic diagnosis is difficult.
Human remains were found at the Aurignacian levels of the Spanish site of El Castillo. Unfortunately, the remains were later lost before a detailed anatomical description could be published. A subsequent assessment by Garralda of available descriptions (1989) suggests that the remains were robust, a trait common to both Neandertals and early modern humans. Other sites such as Hahnöfersand and Vogelherd (Street et al. 2006), once thought to date to the Aurignacian have since been dramatically redated to more recent periods.
Evidence of a modern human-Aurignacian association are somewhat better at the sites of Zlatý kůň in the Czech Republic and Kent's Cavern in England. Perhaps, the best evidence we have comes from the site of Mladeç in the Czech Republic. Bone points, perforated animal teeth and a few lithics have been found there. The assemblage appears to be Aurignacian and is associated with skeletal remains that have been well dated to around 31,000 radiocarbon years.
A recent study by Bailey et al. (2009) attempts to shed further light on the makers of the Aurignacian. Many Aurignacian sites have dental remains but they largely have not been used in taxonomic identification. The authors of this paper use Bayesian statistics to classify individual based on the teeth. They used teeth samples for which taxonomy was known to test the accuracy of their technique. In cross validation of known samples, 89% of both Neandertals and modern humans were correctly classified. In the subsequent analysis of the 34 unknown samples associated with Upper Palaeolithic industries, 29 were assigned to modern humans. This is perhaps the strongest evidence to date that modern humans made the Aurignacian. However, this study cannot completely rule out the possibility that Neandertals could have been responsible, albeit for in small part, for the Aurignacian.
References
Bailey SE, Weaver TD, Hublin J-J. 2009. Who made the Aurignacian and other Upper Paleolithic industries? Journal of Human Evolution.
Garralda MD. 1989. Upper Paleolithic human remains from El Castillo Cave (Santander, Spain). In: Giacobini G, editor. Hominidae: Proceedings of the 2nd International Congress of Human Paleontology. Turin: Jaca Book. pp. 479-482.
Kozłowski JK. 1982. Excavation in the Bacho Kiro Cave (Bulgaria): final report. Warsaw: Panstwowe Wydawnictwo Naukowe.
Smith FH, Trinkaus E, Pettitt PB, Karavanić I, Paunović M. 1999. Direct radiocarbon dates for Vindija G1 and Velika Pećina late Pleistocene hominid remains. Proc Natl Acad Sci USA 96.
Street M, Terberger T, Orschiedt J. 2006. A critical review of the German Paleolithic hominin record. Journal of Human Evolution 51(6).
Above photo by Wessex Archaeology under creative commons license.
View Comments
The origin of bone words
Fri, May 29 2009 09:31 | Linguistics | Permalink

Temporal: Comes from a Latin word meaning a correct mixture or balance in the sense of tempering a metal. It later came to mean a mental balance. It is related to the word temper which we have come to associate with its negative sense, i.e. to lose one's temper.
Sphenoid: Many bones are named for their shape like this one which comes from the Greek sphēnoeidēs meaning wedge-like.
Orbit: The Latin word orbis had the meaning of round or spherical and referred to the shape of the eye sockets.
Nasal: Latin nasus meaning nose.
Nasal concha: Concha is the latin word for a shell or mollusk.
Vomer: This word means ploughshare or stylus in Latin and the bone is so named for its shape.
Lacrimal/Lacrymal: This bone, located in the eye socket, gets its name from the Latin word for tear, lacrima.
Ethmoid: From the Greek word ēthmos meaning sieve.
Zygomatic: Related to the Greek word zugon which mean yoke.
Palatine: The Latin word palatum has the meaning palate or sense of taste.
Hyoid: Named for its shape, this bone was thought to resemble the letter upsilon "υ" in Greek and in fact the word huoeidēs means upsilon-like.
Axial Skeleton
Scapula: Comes from the Latin scapulae "shoulder blades", which in turn is related to the Greek skaphein meaning to "to dig out". It is possible that the shoulder blades were used as digging tools in times past.
Clavicle: The Latin word clavicula has the meaning of small key, the diminutive form of clavis (so called because of its shape). It is a loan word from the Greek kleis which means "key, collarbone."
Rib: Has its roots in the Old English rib, and ribb of Germanic origin and further related to Dutch rib(be) as well as the German Rippe with the meaning of "a covering", from the Proto Indo European root rebh- meaning "roof."
Vertebra: Comes from the Latin vertebra meaning "joint or articulation of the body, joint of the spine", probably from vertere meaning "to turn" with the -bra suffix relating to the arms.
Manubrium: Come from the Latin manus (hand) with the meaning of handle or haft.
Sternum: Came to English through Latin via the Greek word for chest sternon.
Sacrum: From the Latin os sacrum, which itself is a translation of Greek hieron osteon "sacred bone." This name may originate from the fact that the sacrum was often offered up as part of animal sacrifices.
Innominate: Comes from Latin innominatus meaning "nameless."
Pelvis: Has its roots in the Latin word pelvis meaning "basin" (Old Latin peluis), and ultimately from the Proto Indo European root pel- meaning "container."
Arm Bones
Humerus: Related to the Latin word for the upper arm or shoulder umerus.
Its roots have been traced to the Proto Indo European *omesos and to the Sanskrit amsah and Greek omos.
Radius: This word comes to English from Latin and as you may have guessed has something to do with the radius of a circle. The meaning was literally spoke (of a wheel, etc.) or ray.
Ulna: Although the relationship may not be apparent at first glance the words ulna and elbow are etymologically related. The Old English word eln shares an Indo-European root with the Latin ulna. The Old English word for elbow was elnboga which is broken down into eln "forearm" and boga "bend, bow." There is a older Proto Indo-European root *el- meaning "forearm, elbow."
Hand Bones
Scaphoid: Comes from two Greek words eidos or εἶδος meaning “like” or “resembling” and skaphe or σκάφη meaning a boat.
Lunate: From the Latin word lunaris meaning moon. Named for its crescent moon shape.
Triquetral: From Latin triquetrus meaning “three-cornered” or “triangular”.
Pisiform: Named for its resemblance (Latin: forma = shape) to a pea (Latin: pīsum).
Trapezium: Named for its trapezoidal shape. The Greek word trapeza meant “table” and is itself derived from two root words tra- “four” and peza “foot”. The latter word is related to the -pus in octopus and -ped in pedestrian and pedometer.
Trapezoid: See trapezium. Literally table (Greek: trapeza) shaped (Greek: eidos or εἶδος).
Capitate: From the Latin caput meaning “head”.
Hamate: Latin hamatus, from hamus meaning “hook”.
Leg Bones
Femur: Comes from the Latin word femur meaning thighbone.
Tibia: The same word in Latin was used for the shinbone but also had the meaning of "pipe" or "flute," presumably because of it's shape.
Fibula: Comes from the Latin word fibula, which meant "clasp, buckle, brooch" probably because its shape resembled that of a fastener. Etymonline suggests that it is probable related to the Greek perone, which had this meaning. On the other hand, the Oxford English Dictionary points to a possible relationship with the Latin figere meaning "to fix," which in turn can be traced back to a PIE root *dhigw- ("to stick, to fix").
Foot Bones
Calcaneus: Comes from the Latin calx, meaning heel as well as chalk. This in turn derives from the Greek χάλιξ meaning pebble or stone.
Talus: From the Latin talus meaning ankle. It is related to the Latin word taxillus, which was a small die, since dice were originally made from the anklebones of animals.
Navicular: Related to the English word navy. This bone was so named because of its similarity to a boat (navicula in Latin).
Cuneiform: From the Latin cuneus meaning “wedge”.
Cuboid: Named for its square shape.
Last Updated: 29 May, 2009
Above photo modified from original by Aeioux under creative commons license.
Darwinism: The creationist straw man
Wed, May 27 2009 01:10 | Scepticism | Permalink

Outside of intelligent design (i.e., creationism warmed-over), Darwinism is used primarily to refer to the theory of the evolution of species by natural selection, as formulated by Darwin, not to the modern and more complete understanding of evolution. As things tend to be in science, evolutionary theory hasn’t stood still since 1859, when Darwin communicated his ideas to the world in On the origins of species. While natural selection is indeed a principle driving force in evolution, it is not the whole story. Charles Darwin would no doubt be astonished by the such discoveries of genes and DNA, the reworking of evolutionary theory to accommodate evo-devo and the neutral theory of molecular evolution, as well as the plethora of evidence that has confirmed the basic tenets of his original ideas. To use the term Darwinism is an insult to the hard work of the thousands of scientists who have helped refine evolutionary theory. It implies that the wheels of scientific research ground to a halt some 150 years ago and serves to confuse the public’s already poor understanding of evolution.
A more menacing motivation for using the term Darwinism is to portray evolution as just another ideology, consisting of its own set of monolithic doctrines and beliefs. However, science is not based on static beliefs but is rather a ceaselessly self-correcting discipline that evolves in light of new evidence.
By misrepresenting evolution as the archaic, dogmatic ideas of one old man, creationists set up a straw man argument. It always strikes me as curious that they don’t refer to adherents of gravity as Newtonists! Judging from the overwhelming and ever-increasing evidence for evolution, creationists are given little choice but to resort to fallacious arguments.
Above photo modified from original by Colin Purrington under creative commons license.
Argumentum ad hominem
Tue, May 26 2009 02:56 | Scepticism | Permalink

This argument comes in the following forms:
Abusive
The tactic is to portray the opponent as a bad or immoral person, and conclude based on this, that their argument should not be accepted, e.g.
Ann says that anthropological research needs more funding. This is coming from a woman who divorced her husband.
Circumstantial
Involves suggesting that the person making the argument is so doing out of self interest, e.g.
She's an anthropologist. Of course, she's going to say that evolution is true.
Tu quoque (Latin: "you too")
This is commonly heard in political debates. Also referred to as the "hypocrisy argument" or the “you too fallacy”, it involves showing that the opponent's arguments or criticisms apply or have applied to them, e.g.
You said Neandertals and modern humans interbreed but only last year you said that they didn't.
Poisoning the well
Is a pre-emptive attack meant to discredit one's opponent before they even make their argument, e.g.
Darwinists have been preaching the evolution lie for years. I suspect that tonight's debate won't be any different.
Guilt by association
Is the view that an individual shares the qualities of others based on irrelevant associations, e.g.
Craniometrics was used in the early twenthieth century to promote eugenics. Anthropologists who carry out craniometric analyses are like Nazis.
Above photo modified from original by Caro's Lines under creative commons license.
Race: an anthropological perspective
Sat, May 23 2009 08:38 | Biological Anthropology, Cultural Anthropology | Permalink

Alice Brues defined race as "a division of a species which differs from other divisions by the frequency with which certain hereditary traits appear among its members." This definition of race, like most others, is rather equivocal, in that it does not tell us how much variance in the frequency of traits necessitates the creation of a new race. If we take this definition at face value then according to craniometric and genetic data an incalculable number of races exist.
In 1972, R.C. Lewontin reported that, for genes at a single locus, most genetic variation existed within populations, rather than between them. For most biologists this put the nail in the coffin for the race concept. In a re-examination of Lewontin's findings, the Cambridge statistician A.W.F. Edwards, noted that our ability to correctly classify populations is due to the correlations among different loci. By focusing on multiple loci the between population differences increase dramatically.
As way of an analogy, imagine we asked a stranger the following question via internet: "Which colour do you prefer less: orange or brown?" Previous surveys have showed that there is a slightly greater tendency for women to rate orange as their least favourite colour, while men have a slightly greater tendency to dislike brown. However, there is a very high amount of overlap. We would have a tough time trying to predict sex based on the results of this single question. However, if we ask say thirty questions instead of one, we would be able to predict sex with a much higher degree of certainty based on the responses as a whole. In a similar manner, we could not confidently determine race of an individual based on one or two cranial measurements. However, the likelihood of a positive determination increases significantly when we include more measurements.
Craniometrics has been shown to correctly classify individuals into a few broadly defined racial categories, as well as many more geographically localised categories. The ability of forensic anthropologists to accurately classify individuals into predefined groups does not substantiate the biological race concept. Just because we can determine a skeleton to be of Irish, Western European, Northwestern European or European ancestry does not mean that such ancestral groups exist in any meaningful biological sense. However, such information is useful for homicide investigators who are interested in whittling down their list of possible missing persons.
So how are we to understand race? Human variation is probably best understood in terms of both temporal and geographic distances. Cranial variation correlates strongly with geography; meaning that the further apart the populations are geographically, the more dissimilar they are phenotypically. Conversely, neighbouring populations show greater phenotypic similarities, spurring anthropologist Frank Livingstone to write in 1962 "there are no races, only clines”. The relationship between phenotypic variation and geography is likely due to both isolation by distance (there is greater gene flow between neighbouring populations) and the many founder effects that occurred in the course of human history. The longer groups remain isolated the more dissimilar their genotypes. Since most racial categories are defined by geographic regions, it should not come as a surprise that there is a correlation between race and place of ancestry.
Race is a crude sociocultural construct based on the underlying reality of biological variation. In this regard it is similar to other cultural phenomena, which help us understand our past. For example, much can be ascertained about ancestry and human migration by studying languages. In this regard, race has proved to be a useful concept in the fields of medicine and law enforcement. As long as law enforcement continue to use racially defined categories, forensic anthropologists will similarly follow suit.
Above photo modified from original by indianfilipino under creative commons license.
A little human with very big feet
Wed, May 13 2009 02:50 | Palaeoanthropology | Permalink

The pelvis and legs all clearly demonstrate that Homo floresiensis was bipedal. However, the hominin’s feet are unusually long compared to the leg. This combination of a long foot and a relatively short leg is seen in some apes but not in hominins. The navicular acts like the keystone in the arch of the human foot and is elevated from the ground except in people who have fallen arches. In Homo floresiensis this bone has a well developed tuberosity meaning that it was in contact with the ground, like in other flat-footed great apes and early hominins. The overall shape of the foot means that this hominin would not have been able to run long distances very efficiently — a distinguishing feature of later Homo. While the hominin had a short chimp-like big toe, it was not opposable like in other hominins. The long, curved lateral toes resemble a chimpanzee’s, rather than those of a human which are short and straight. The distal first metatarsal is squared off like in modern humans but this feature is not found in other early hominins, such as Australopithecus afarensis, Paranthropus robustus or the early Homo remains from the Georgian site of Dmanisi.
It is generally thought that Homo erectus was the first hominin to leave Africa, soon after their first appearance in the archaeological record around 1.9 million years ago. Based solely on archaeological data Homo erectus seems like the best ancestor for Homo floresiensis. Homo erectus has been show to be a highly variable species. Homo floresiensis could represent a descendent of Homo erectus that adapted to island life through a dramatic decrease in size. While some of the plesiomorphic traits of Homo floresiensis may be explained through evolutionary reversals, it is unlikely to account for all of the primitive traits in the skeleton as a whole. It has been suggested that the ancestor of Homo floresiensis was not Homo erectus but rather a more primitive hominin. The 1.8 million year old skeletal Homo remains from Dmanisi are relatively primitive. At first glance, this species might seem like a good ancestral candidate for Homo floresiensis. However, unlike Homo floresiensis, the Dmanisi specimens have quite modern limb proportions. Homo habilis has also been forwarded as a possible candidate although there is little archaeological evidence to suggest that that species ever left Africa. Until further evidence comes along, the jury is out on this miniature human.
Reference cited
Jungers et al. The foot of Homo floresiensis. Nature 459, 81-84 (7 May 2009) | doi:10.1038/nature07989
Above photo modified from original by Mamoritai under creative commons license.
Is craniometry scientific?
Wed, May 13 2009 02:07 | Biological Anthropology, Scepticism | Permalink
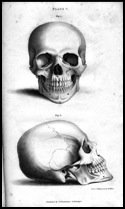
Craniometric studies were, to a large degree, racially motivated in the early decades of the twentieth century, with anthropologists trying to validate their preconceived racial categories. In 1912, Franz Boas published a study challenging the prevailing notion that certain cranial measurements were under ironclad genetic control. He studied the head form of some 13,000 European immigrants and their American-born children. He found significant differences in the shape of the heads between parents and their children, which he interpreted as evidence for cranial plasticity. In other words, environment, not genetics, shapes cranial morphology. During the subsequent decades, Boas' results came to be largely accepted by the anthropological community, with students of the Boasian school disregarding craniometric studies as an ill-fated enterprise. For many years, anthropologists steered clear of craniometry, instead focusing their expertise in other less stigmatised areas of physical anthropology such as palaeopathology.
Quite recently papers by Sparks et al (2002) and Gravlee et al (2003) have re-examined Boas' original data. However, those expecting the final word on cranial plasticity were to be disappointed. The anthropologist Milford Wolpoff is quoted as saying in 1975 "The data do not speak for themselves. I have been in rooms with data and listened very carefully. They never said a word." This is particularly true of these two papers, which use the very same data to come to divergent conclusion. While Gravlee et al believe Boas to be essential correct, Sparks et al came down firmly on the other side. The real answer, I believe, is to be found between the lines.
Boas did, in fact, find a statistically significant environmental effect in his study but this begs the question of whether it is a meaningful effect. Sparks suggests that while the effect is real, it only constitutes a tiny proportion of variation. In fact, considering the size of Boas' sample (~13,000) it is almost impossible not to find statistically significant results; biology is, after all, intrinsically variable. It may be the case that Boas played up the importance of the environmental effects as a reaction to the racial thinking that was prevalent at that time.
Perhaps, the biggest problem with Boas' methodology was his reliance on only a handful of measurements and particularly the use of the cephalic index (ratio of head breadth to head length). Anders Retzius introduced the cephalic index as way of classifying skulls based on their overall shape. He defined three main categories: dolichocephalic (long headed), brachycephalic (broad headed) and mesocephalic (intermediate headed).
Most modern biological anthropologists are of the opinion that the use of a couple of measurements to describe a multi-complex structure such as the skull is absurd. Today, biological anthropologist will take dozens of measurements of the skull. W. W. Howells, who measured thousands of skulls from all over the world, had the following to say about the cephalc index: "When Anders Retzius, a century and half ago, invented the cranial index, he gave us an answer for which there was no question." Even Boas himself wrote the following in 1940: "Measurements should always have a biological significance. As soon as they lose their significance they lose also their descriptive value."
Craniometry is used today in biological anthropology as a means of determining the relationships of peoples through their phenotype. A phenotype is the visible manifestation of a genotype. Since there is rarely a one to one relationship between the genotype and phenotype we must first demonstrate that the phenotype is an accurate reflection of the genotype. If this is not the case craniometry would be no more scientific than phrenology. Narrow-sense heritability is the proportion of phenotypic variation that arises from only the additive genetic differences among individuals and is expressed as h2 = VA/VP. Heribtaility is measured on a scale of 0 (no heritable variation) to 1 (all phenotypic variation is due to additive genetic effects). The average cranial h2 has been estimated at around 0.55 (Relethford 1994; Devor 1987). A heritability greater than 0.5 indicates that most phenotypic variation is the examined traits are attributable to genetic factors. Thus the proportionality of genotypic to phenotypic variance is a reasonable assumption. The true litmus test of any hypothesis is its predictive power. Craniometric data is used with surprising accuracy by forensic anthropologists to determine likely ancestry of unknown individual and by palaeoanthropologists to determine our relationship to other hominins. The modern scientific practice of craniometry distinguishes itself from psuedosciences like phrenology and physiognomy in that it is based on sound biological theory, it is testable, it is predictive and objective.
References cited
Boas F (1912) Changes in the bodily form of descendants of immigrants. American Anthropologist 14: 530-562.
Sparks CS, Jantz RL (2002) A reassessment of human cranial plasticity: Boas revisited. Proc. Natl. Acad. Sci. USA 99: 14636-14639.
Gravlee CC, Bernard HR, Leonard WR (2003) Heredity, Environment, and Cranial Form: A Reanalysis of Boas's Immigrant Data. American Anthropologist.
Devor EJ (1987) Transmission of human craniofacial dimensions. J Craniofac Genet Dev Biol 7: 95-106.
Relethford JH (1994) Craniometric variation among modern human populations. Am J Phys Anthropol 95: 53-62.
Ad Hominin. What's in a name?
Wed, May 13 2009 02:04 | Scepticism | Permalink
Until quite recently humans and their ancestors were referred to as hominids. This was a reflection of taxonomy of the time: the family Hominidae consisted of humans, while the family Pongidae consisted of orangutans, chimpanzees, gorillas and bonobos. However, molecular studies showed humans, chimpanzees and bonobos to be more closely related than previously thought. The current consensus taxonomy for the family Hominidae consists of the following subfamilies: Ponginae (orangutans), Gorillinae (gorillas) and Homininae (chimpanzees, bonobos, humans). Strictly speaking chimps, bonobos and humans are all hominids. However, we still need a way to talk about ourselves and our ancestors. The Homininae subfamily is divided into two tribes: Panini (chimpanzee, bonobo) and Hominini (humans). Thus, in recent years there has been a growing trend of referring to humans and their ancestors as hominins.The Latin prefix ad means "towards", so ad hominin can be translated as "towards humankind". Many of you will undoubtedly notice the allusion to argumentum ad hominem - a type of argument where one attacks the character of the opponent rather than their arguments. This pun is intended and reflects my interest in critical thinking and logic.Many people lack the cognitive toolkit to critically examine such psuedoscientific claims as homeopathy, psychokinesis or acupuncture. As a physical anthropologist I am well aware that evolution is not impervious to wild misconceptions. It is my hope to give people the tools and knowledge necessary to recognise fallacious arguments and critically examine extraordinary claims.